
Ring-porous

European Ash (Fraxinus excelsior) is an example of a ring-porous wood, with the earlywood pores clearly forming rings or bands—in this case two to four rows wide.
Diffuse-porous

Afzelia (Afzelia spp.) is an example of a diffuse-porous wood, with no clear earlywood-latewood pore arrangement, and no significant difference in pore size.
Semi-ring-porous

Butternut (Juglans cinerea) is an example of a semi-ring-porous wood. Although the pores don’t form clear rows, the size gradually decreases from the earlywood to the latewood.
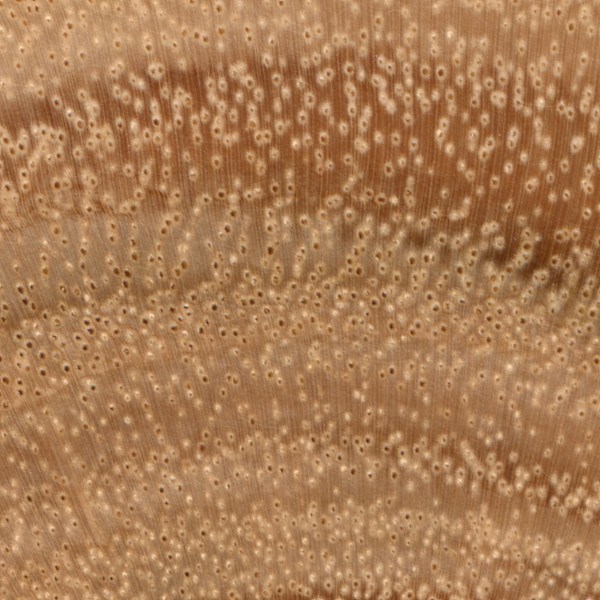







Pore contents
Besides simply considering the size or arrangement of the pores, sometimes it pays to observe what’s actually in them. As sapwood becomes heartwood, certain substances and structures are deposited in the wood cells.
In some cases, pores can become filled with colored gums, resins, or other deposits that can aid in identification. In most species, heartwood deposits tend to be somewhat sporadic, so it shouldn’t be relied upon as a primary identifying feature. The frequency of pore contents, if present, are described using the relative terms, sparse, common, and abundant.
Another common by-product of the conversion of sapwood to heartwood is the appearance of tyloses. Tyloses (singular, tylosis) appear as bubble-like structures that grow into open pores, and in some cases, completely stop-up the pores of the heartwood.
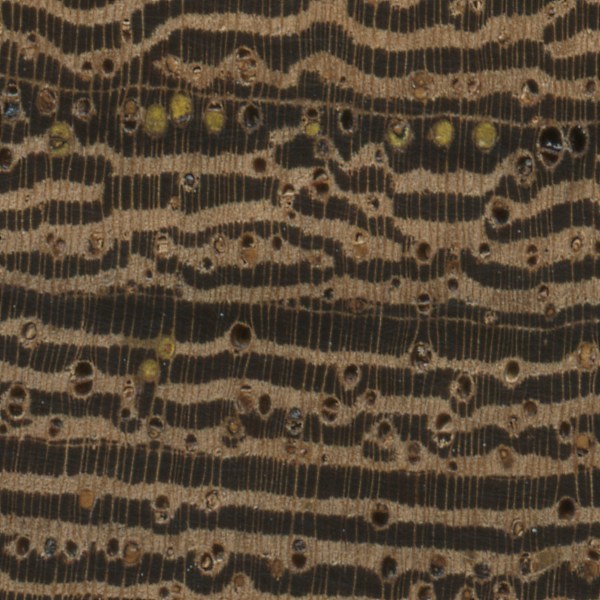
The sample of Panga Panga (Millettia stuhlmannii) has quite a few heartwood deposits; many are a striking yellow, while some are a dark brown.

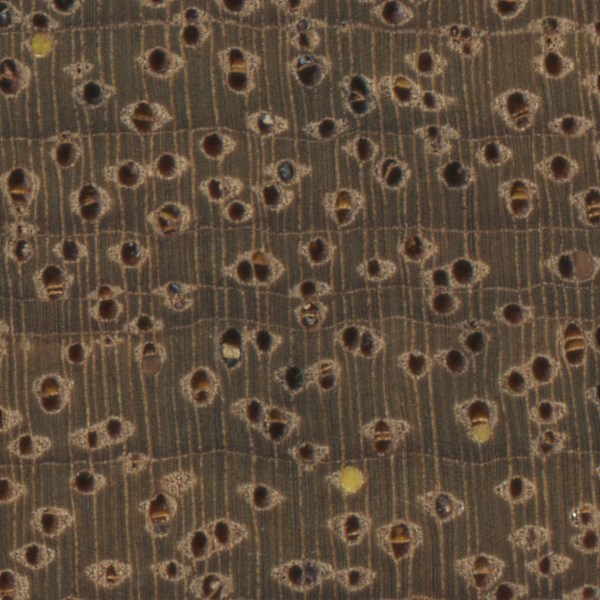
Black locust (Robinia pseudoacacia) is a superb example of a wood with pores that are abundantly packed with tyloses.
Some species lack tyloses altogether, while many others have an intermediate distribution ranging from sparse to common, while some species have tyloses in abundance, to the point that nearly every heartwood pore is filled with tyloses. This blockage has the beneficial effect of rendering the pores essentially watertight: a well known example of this is found in White oak (Quercus alba), whose packed pores are commonly used for barrel-making (also known as cooperage).
Apotracheal parenchyma
In order to understand this somewhat intimidating scientific term, it may help to consider the English word “trachea,” which refers to a tube or pipe (in this case, a wood pore). Combine this with the Greek prefix “apo,” which means away from or separate, and the meaning becomes clearer.
Apotracheal refers to parenchyma cells that occur separate from the pores. Apotracheal parenchyma can occur as single scattered cells, classified as diffuse parenchyma. These cells are too small to be seen without a microscope. However, in some wood species, several apotracheal parenchyma cells are joined or aggregated together, forming thin but visible tangential lines. This formation is known as diffuse-in-aggregates parenchyma.

Honduran rosewood (Dalbergia stevensonii) displays a multitude of diffuse-in-aggregates parenchyma, typical of many Dalbergia-genus rosewoods. Any apotracheal parenchyma in contact with neighboring pores is incidental.












Aggregate rays
Occasionally, some species will have intermittent rays that are many times wider than the rest. These mega-rays are essentially a collection of a number of normal-sized rays grouped together and appearing as one large ray. They are known as aggregate rays.
Perhaps the most well-known commercial lumber in the United States that features aggregate rays is red alder (Alnus rubra). In addition to alder, only a handful of other genera and species exhibit these rays: hornbeam (Carpinus spp.), and some species of sheoak (Allocasuarina and Casuarina spp.) and oak (Quercus spp.) also feature aggregate rays.

Occurrence of the large rays is very sporadic—some samples may not contain any aggregate rays at all, while other pieces will have several. The aggregate rays of red alder are so large and conspicuous that they can be observed on flatsawn surfaces as thin dark streaks, and may be mistaken for defects in the wood.
Noded rays
In some species, the rays will slightly flare out and get wider as they cross a growth ring boundary. This subtle characteristic is referred to as noded rays.
While noded rays are fairly uncommon, they can occasionally serve to help in identifying certain woods. Noded rays are present in sycamore (Platanus spp.), beech (Fagus spp.), basswood (Tilia spp.), and yellow poplar (Liriodendron tulipifera).

Storied rays
One final characteristic of rays involves examining the flatsawn (tangential) surface of the wood. In some wood species, (particularly those in tropical regions), the rays tend to be aligned in horizontal or diagonal tiers, also referred to as stories. This pattern is called storied rays, and it produces a visual phenomenon known as ripple marks.Even though there’s technically no unevenness in the wood, to the unaided eye, (and at low levels of magnification), storied rays appear as minute stripes of wood alternating between light and dark. In addition to the rays, other anatomical features (such as the parenchyma or the fibers) can also form stories, and therefore produce ripple marks.

In addition to persimmon (Diospyros virginiana)—a temperate species—some notable tropical species with storied structures include: most rosewoods (Dalbergia spp.), Honduran mahogany (Swietenia macrophylla), and sapele (Entandrophragma cylindricum).




Bamboo
These fast growing members of the grass family (Poaceae) are very diverse, with hundreds of species encompassing dozens of genera. Unlike trees, bamboo initially grows at full width, with no tapering or horizontal growth.
Much like palm, bamboo has fibrovascular bundles that are more concentrated near the edge of the stem—also called the culm—with softer parenchyma grading in toward the inner wall. Bamboo culms are hollow in the center, with closed “nodes” generally occurring every few feet or so.


The sample above is Bamboo (possibly Phyllostachys spp.), which shows subtle interruptions in the grain pattern at the nodes (seen in the right-hand third of the face grain portion). The endgrain also shows smaller strips that have been laminated to form a larger plank.

Note the increasing concentration of fibrovascular bundles near the edge of the endgrain section of Giant Bamboo (Dendrocalamus asper).
Because bamboo has such a small (and hollow) diameter, various techniques are used to manufacture what could be loosely termed as “boards.” Uniformly small strips of bamboo and machined and then glued together to form larger planks, slabs, and plywood for a number of applications, including, flooring, construction, and furniture.
Get the hard copy
 If you’re interested in getting all that makes The Wood Database unique distilled into a single, real-world resource, there’s the book that’s based on the website—the Amazon.com best-seller, WOOD! Identifying and Using Hundreds of Woods Worldwide. It contains many of the most popular articles found on this website, as well as hundreds of wood profiles—laid out with the same clarity and convenience of the website—packaged in a shop-friendly hardcover book.
If you’re interested in getting all that makes The Wood Database unique distilled into a single, real-world resource, there’s the book that’s based on the website—the Amazon.com best-seller, WOOD! Identifying and Using Hundreds of Woods Worldwide. It contains many of the most popular articles found on this website, as well as hundreds of wood profiles—laid out with the same clarity and convenience of the website—packaged in a shop-friendly hardcover book.
Could anyone at all help me identifying this wood? It’s a circular frame, 19cm diameter.
Any recommendations on wood identification for someone with trypophobia? I very much appreciate this website and all the work that Eric has put into this, but I can only come here on ‘good’ days.
Having a real tough time identifying this one.
These are wood types I have for comparison. Interested in the purple heart. Grain is sappy and sticky. It is aged stored upright on end with some twist and warp. Was told it is irritating And to cover one’s self when milling. Can you add any explanation and milling safely to follow?
Hi Eric,
I came across a 14″ log (crotch piece) in a random pond and couldn’t resist turning it. No smell, generally cut like butter, and is relatively light. Any ideas? Thanks
Pics are: turned bowl, iPhone macro, 10x, 20x.
I’m guessing probably something on this page: https://www.wood-database.com/wood-articles/poplar-cottonwood-and-aspen-whats-what/ Some of these woods can be VERY hard to tell apart though.
Thanks for taking the time to respond, Eric. I appreciate it. Certainly plenty of poplar and cottonwood around here (Williamsburg, VA). Was thinking it might be Black Tupelo, but poplar would make sense too given where I found it.
Tilia sp? What little I see of the bark it looks thick and fibrous. In mature Tilia bark (multiple mm thick), there would be a distinct, tough net-like layer that can be easily separated. It’s a very light and soft wood, easy to shape. I’ve made a mounting bolt out of it, by simply twisting a slightly oversized piece into the corresponding nut. Populus bark AFAIK is much thinner.
Hello again!
I am posting another 2 samples of a species and I was wondering if these could be genus Dalbergia (Palissander, Rosewood), Teak or another genus.
Thanks again for any help!
Hello!!
Congratulation for the article and thank you for the informaiton.
If you have any idea of this 2 species?
Could it be genus Dalbergia (Palissander, Rosewood) or Teak?
Thank you in advance.
haloo?? have any idea what their names species
Have a look at this page: https://www.wood-database.com/locust-osage-and-mulberry/
Here is the face grain
Anybody have a clue I was giving this out of a really old barn and I believe it was a post at some point in time It’s very dense and I live in northeast Tennessee
Looks like black locust to me.
Hello and thank you for this good article.
Besides the identification process, does each of these characteristics have an impact on the object we can create with the wood ?
I read your article here which covers a little what I’m asking: https://www.wood-database.com/wood-grain-texture/
If you have any other article which covers more the impact of the wood characteristics on the creation, I would be glad to read it.
Thanks again !
I would say in general, no, the individual anatomy of the wood species don’t seem to have any strong correlation between other characteristics like strength or stability. But I think musical instrument makers have bounced around a lot of theories and ideas of what characteristics go into making a good tonewood, but that is perhaps another discussion.
Yes, anatomical characteristics influence all wood properties, and so do ecological variations within the same species. That is why knowledge about wood anatomy is so important for all activities that use wood as a raw material.
There are numerous articles showing these correlations to different propositus.
Hello Eric,
I’m curious how you get such wonderful photos of the endgrain; obviously need a good scope/camera, but is there any trick to preparing the specimen? I have tried sanding, planing, and sawing but the pores seem to get clogged up or mangled by the tool. Thanks!
There’s two schools of thought on this: the microscope folks and the folks that just sand the wood. I’m in the second camp. It looks like you are cutting the wood from the linear cut lines and rough edges, sort of what a microtome would do to prepare a microscope slide. I just finely sand the endgrain up to about 800 grit, then switch to the foam-backed sanding pads such as Abralon at 1000 grit (I think Festool makes one just like Mirka’s product too). I’ll sometimes go up to 2000 grit for darker woods with the Abralon. The cushioned… Read more »
Surprising to hear from Eric that sanding is actually effective. I did not try sanding while preparing one specimen for identification, assuming that it would not work. What did work, after verifying that my plane irons were sharp and still getting lackluster results, was using a sharp chisel. Bevel-up of course, sometimes having to lift a few degrees to get it to actually bite.
My pics got mixed up, that’s actually the upper-right of the plank, I.e. the 2 boards, etc. on the right. Hopefully, you can still make out the delineation of boards, ring boundaries, and pores. Thanks.
Pulled these 4-board plank off a piano (where it was under a veneer and substrate). I can’t tell whether it’s diffuse pores or semi-diffuse/ring, such that it might indicate the type of wood (e.g. maple/beech/birch or walnut, respectively). It seems clear it’s not ring pores. Any thoughts on the type of wood? Pictured here is a shot of the left 2 boards. One from 1-ft away, showing the 2 boards and plainly showing the ring boundaries; and, the other pic is a closer shot of the distribution of pores on the right board. ‘Love the site. Thanks for all the… Read more »
Can you get a picture of the endgrain? Either sand it or take a thin slice off with a miter saw to clean up the end, then snap a pic.
Thanks, Eric. I had to cut off the end, and sand (applied boiled linseed oil, too). Here’s a couple pics, pre-sand and post-sanding. That raw ‘soft’ wood appearance (and, greener looking) now makes me think poplar – and, it’s not a heavy wood like maple, etc. Thanks for forcing me through a better identification process/assessment. But, please….. let me know if I’m wrong. PS: I’ve deconstructed 5 upright pianos over 100 hrs old each, and poplar was a well-used wood – whether as a substrate to some nice veneers or as a block-laminate wood. So, it wouldn’t surprise me to… Read more »
I agree that the ray fleck seen in the previous photos is also reminiscent of poplar, so it seems a very likely candidate. The last two photos are a ring porous hardwood of some sort, but they don’t appear to be oak. Oak has very wide and conspicuous rays, and I can’t make out any sizeable rays in your photos. Possibly ash or elm?
Thanks Eric. Poplar seems reasonable, i.e. color, weight, “soft” cut, distribution of diffuse pore with rays throughout. The other plank wood definitely appears to be a ring porous wood, but I am easily fooled sometime by the semi-ring porous woods, i.e. ask, to me, seems to have distribution of pores well into the early wood growth; or, true hickory woods, for example, also seem more alike semi-ring porous although defined as ring-porous. Anyway… thanks again. We very much enjoy the site.
I would take a look at what you have compared to Quartersawn Sycamore. Not only it looks very similar and has the same checkerboard pattern and is very common to be mistaken with maple.
Hi eric
its a great website
i wonder if you could help me to identify this wood based on these photos
it really hard, so heavy, kinda mixed redbrown color
maybe it is similar with some kind of Ipe, Massaraduba, Cumaru or else.
Thanks
Those darker black streaks in the second picture remind me more of curupay (aka cebil). Otherwise goncalo alves can also be streaked like that. Getting a clear, finely sanded closeup picture of the endgrain might help.
This looks very reminiscent of old aged purpleheart (the wood becomes a dark muted purple like in the images over time) the grain patterns also looks very similar to a lot of the purpleheart ive worked with. If you cut the wood or sand it deeply, then wet it and expose it to the sun for a day or two, it will likely turn a bright purple.
I am thinking to purchase some french doors, but the seller is not able to identify the type of wood. I am looking for oak. Are you able to confirm if this is oak, or give me clues to what to look for to tell? Thank you,
Hi eric,
I’ve been trying to identify this wood I know it came from a sawmill where they make rocking chairs it is very hard and heavy it has been seasoned but the thing I like about most is the color. Can you please help?
Please clean up the endgrain and expose fresh surface and repost photos. The surface is just too weathered to make out any details.
Thank you for your support but recently I ran into a friend of mine who has a sawmill and he informed me that it was actually poplar wood that I have. He explained to me that some of the wood is only harder and heavier because it has absorbed different minerals from the ground causing the beautiful colors and in turn making the woods so hard to work with.
Do you have any suggestions on working with this type of hardwood?
Poplar is very easy to work, the only problems you might have would be if you wanted to stain it a certain color instead of leaving it natural color. It can absorb stain unevenly and cause blotches, so you’d probably want to look for a gel stain.
Hey Eric!
Your website has become invaluable to me, as I use it all the time when working with different types of woods and just to browse through your articles.
I was wondering if you could help me figure out what this piece of wood is, it has me completely baffled. I received a few planks that are about 3 feet long, 3 inches wide, and half inch height.
Thank you for all the hard work you put into this website and helping others!
Darren
Well, it looks like a diffuse porous tropical hardwood of some sort. Those wide rays look really distinct, and remind me of Cordia species, but that’s just a guess. https://www.wood-database.com/wood-filter/?fwp_genus=cordia
Ipe is my guess
Hi Eric, thanks for your article! I It’s really helpful. Recently, I have been studying on grains as the inspiration for my design, but there is a puzzle I couldn’t solve even after searching for many resources…Do you know how this formed? Why the late wood is elongated with many intervals of early wood? This is a ring-porous wood. Is there something to do with pores? Also, as for the second page, round grains are formed around knots. But why there is a clear boundary between late wood and early wood not early wood to late wood as on the… Read more »
I’m sorry, I don’t understand what you are asking. I can’t see what you are referring to and circled in red. Many ring porous woods form rows that are multiple pores wide, so that may be what you are seeing?
The abruptness of the grain is mostly due to the growth rates of the tree through the seasons. Since it gradually slows down and stops in the fall and into winter, and then abruptly starts back up again in the spring, this is reflected in the wood too.
Hi Eric, This is a great resource. I’m trying to determine what family of wood this sample is from. I’, interested in making a guitar so I’m looking for a stable fairly dense timber similar to Mahogany or Queensland Maple (Australia). I have a few good pieces of this timber I was hoping to use.
What is the weight of the wood? Any noticeable scent when working it? Can you post facegrain pics? Where did it come from?
Great article! Thanks a lot for the documentation.
Does the less pores from Brazillian rosewood has something to do with sound vibration? I guess this is one of the main reason why BR is sought after in musical instruments!
Hi Eric, here is a very clear magnification of a petrified wood specimen, the real piece is only 25 mm by 15 mm. I expect its a piece from Chinchilla, Queensland, Australia. Can you ID the wood? Im not an academic, just an interested person. Its a macro photo with a Samsung A71
I’m afraid I don’t have a chance with fossil wood like this. You might try using NCSU’s Inside Wood system for fossil woods. Maybe you could even contact them and see what they think. https://insidewood.lib.ncsu.edu/search?0
Ok, thanks Eric, appreciated you had a look at it. Ive got no chance at all :-))
Do you have your book for sale I could buy? Amazon currently do not post to Australia, and the bookshops here in Canberra do not have it, and say they cannot get it.
I don’t think it’s available directly to Australia at the moment without using some sort of package forwarding service. Sorry!
Hi Girts.
If it is from the Chinchilla area then it might be worth comparing it to Eric’s description for Brigalow (Acacia Harpophylla) – https://www.wood-database.com/brigalow/
Would these pores be solitary, and winged, narrow rays?
Yes, the pores are exclusively solitary, with winged parenchyma. Rays may be closer to medium width, hard to say. At the moment, there’s no matches for those characteristics on the Wood Database. The most common genus with solitary pores is Eucalyptus, but winged parenchyma throws a wrench into the gears.
any idea what wood this may be? this sample is fossil wood, no idea from where, expect its Australia
A fossilized Australian wood. I have about a 0% chance of getting this one! Sorry!
ok, thank you!
I am trying to identify the wood on my table and chairs. It is very hard and looks like it has eyes in several places.
Do you know where I can access to a database of end-grain photos for each type of wood (20-50 samples)?
You can try here: https://insidewood.lib.ncsu.edu/ Definitely not going to get 20-50 though, maybe 5-10 is more reasonable. And they are microscope images, not photos per se.
Can someone tell me what this wood is from the end-grain photos?
Sweetgum or maple or hackberry?
3rd pic
Second pic.
I live in Los Angeles and my neighbors were cutting down a tree so I saved a few horizontal slices (like tree cookies) but I’m not well versed enough in the myriad of wood species to identify it. I have pictures of the unfinished wood with some bark on it still and some of it sanded but un-treated still. I can’t find photos of the tree or its leaves though…. If you recognize it and can let me know what it is, I would greatly appreciate it!!
I have a piece of wood that looks just like this – it is from an Allspice tree – I bought it in Hawaii. Mine does not have the bark, so I can’t compare that.
At first glance I thought this was mahogany, but after looking further, I am thinking walnut. What do you think? I would appreciate your thoughts. Thank you; this is an awesome website.
That definitely shows many of the characteristics of walnut, and not so much mahogany. Since walnut goes blonde as it ages it would make sense that the color of an old chair that was originally either stained reddish-brown or finished with ruby or garnet shellac would exhibit more of the red and less of the brown as it ages.
It is hard to tell conclusively, but I would argue that it has more characteristics of mahogany than walnut. The back rest piece has very tall/long pores that are evenly spaced. Black walnut tends to have much shorter pore lines (perhaps 1/4″?) when viewed on the face grain, while mahogany can have longer pore openings of around 1″.
Found a Henrendon tabletop part piece in trash. Wondering what this wood is. Pretty hard. Had veneer on it.
Hello!! This was found in an old house in Missouri. For the life of me I can’t identify it. It looks like it may have been a flooring piece.
Is it fairly heavy and hard? If so, it’s probably leopardwood.
Hi Eric,
I love your site and use it all the time. What do you use to get the 10x magnification on end-grain? is there a reasonably priced device/microscope you could recommend. I dont seem to be having success with a phone or cheap microscope.
Regards
Brendan
Yes, you can do a search for a jeweler’s loupe. There’s a picture/link to one towards the end of my article on wood identification: https://www.wood-database.com/wood-articles/wood-identification-guide/
Does anyone know what wood this is it is very hard
Sapele?????
The color looks pretty light for sapele. Can you try another pic, this one doesn’t seem to be in focus.
Endgrain pic. Pardon the sanding scratches.
To me, the endgrain is a dead ringer for maple. The bark is somewhat confounding, but there’s a lot of variation across maple trees in terms of bark. Does the wood have much of a scent when being worked? If you’ve worked with cherry before, the faint odor should be recognizable.
Thanks Eric, I couldnt find my post! Thanks for the reply. No I havent worked this wood yet. i have worked with cherry but as you say the endgrain is different. Thanks.
Found this petrified wood in oregon at work. It has tight zigzag growth rings, any idea what type of tree it came from?
Great resources! Ive been studying all available to ID a couple boards I have and what I came up with is maybe wild cherry. The bark says cherry but the endgrain could be cherry or maple. Ive included a couple pics. Am I close?
What kind d of wood is this ?? It is very dense and heavy. Feels waxy after cutting and tried sanding and it barely made a difference. I hit it with a hammer it it made just a small divot..
Can’t tell from the pic, sorry. It appears to be spalted sapwood of some sort. That little bit of dark heartwood in the lower right looks interesting. Doesn’t seem to be your run of the mill stuff.
and here’s a front piece
Here is the end grain
Anyone have any idea what this wood is? It is a vanity that came from an old lodge about 40 years ago. It is heavily stained, can see a nice grain and knots. Will add pics of the end grain and a stripped part.
dear sir,
iam also the wood anatomy experience holder person, trained from FRI,
Dehradun. Iam highly impressed by the deep clearity of the wooden structures. This can be great understanding of wooden species in general.
thanks
rajesh
This is beautiful. I’m guessing Spanish Cedar or Honduran Mahogany. From a garage in Oklahoma. Very lightweight. I can’t pick out a smell. What do you think or know?
Philip,
You can try NCSU’s service:
http://insidewood.lib.ncsu.edu/menu/type/modern/.1
I have a sample of wood from the Blue Mtns near Springwood NSW from a fallen tree. I would like to identify the species. It is red i colour and has small pores without rings and thin <5mm sap. How can I go about this? I have a 10x loop. can i key this out using a series of micro-graphs and working backwards from a list of tree species in the area? Can you advise links? Many thanks Phillip
Hello, I make outdoor hammocks made from wooden dowels. Originally I was using asian birch and treated them with 3 coats of Sikkens HLS (Cetol 1 in North America). This seems to have worked well if there is a re-coat every 2 or 3 years. I am now living in Indonesia, and I want to use teak dowels. Im trying to use scrap pieces from other manufacturers to make the dowel, so they cannot guarantee me premium grade teak. As long as it’s dried 8% – 12% do you think I would have a problem with splitting or cracking with… Read more »